

Cours de SVTHEEB Premiere D – L’énergétique biologique
Objectifs :
• Relever que la dépense énergétique d’un organisme est permanente et varie selon les conditions du milieu
• Définir la notion métabolisme de base d’un individu ou métabolisme basale
• Expliquer les mécanismes de la respiration et des fermentations
• Définir les termes quotient respiratoire et intensité respiratoire
• Calculer le quotient respiratoire et l’intensité respiratoire
• Relever que l’ATP est l’unique source d’énergie directement utilisable par les cellules
• Comparer le rendement énergétique de la respiration et de la fermentation
• Expliquer les mécanismes de biochimiques de synthèse de l’ATP dans les cellules.
I. La permanence de la dépense énergétique et sa variation
1) Permanence de la dépense énergétique
L’organisme dépense de l’énergie de façon permanente sous forme d’énergie mécanique (travail musculaire) et sous forme de chaleur (chaleur rayonné par le corps dans le milieu extérieur ou chaleur sensible notée \(S\) et la chaleur de vaporisation de l’eau dans les poumons et sur la peau ou chaleur latente noté \(L\). Les besoins énergétiques sont évalués soit d’après la dépense d’énergie : on parle de calorimétrie physiologique directe ; soit d’après la production d’énergie : on parle de calorimétrie physiologique indirecte
a) La calorimétrie physiologique directe
Elle s’emploie chez les petits mammifères. On les enferme dans une enceinte et on mesure la quantité de chaleur dégagée dans le milieu ambiant.
On construit des calorimètres à paroi isolante afin d’éviter les contacts avec les milieux extérieurs. Dans la chambre calorimétrique, la chaleur perdue par le sujet est recueillie par un radiateur à ailette dans lequel circule de l’eau froide. Un dispositif permet de régler le débit de cette eau de façon que la température de la chambre reste constante. Connaissant la masse d’eau en Kg, la température \(T_1\) de l’eau à l’entrée et la température \(T_2\) à la sortie ; la déperdition de chaleur sensible est donnée par la relation :
\(S = M({T_2} – {T_1})\)
La Chaleur latente est la chaleur échangée avec le milieu extérieur lors d’un changement d’état : solidification, fusion, ébullition…
Le renouvellement de l’atmosphère de la chambre est assuré par un circuit comprenant :
• Une pompe
• Un dispositif d’alimentation en oxygène
• Un flacon de potasse fixant le \(C{O_2}\)
• Un flacon d’acide sulfurique fixant la vapeur d’eau.
Si la masse du flacon contenant l’acide sulfurique augmente de « m grammes » à une température de 15°C, on peut calculer la chaleur latente. A 15°C, l’évaporation de 1L d’eau nécessite 580 Kcal. La chaleur latente est donc \(L =0,58 \times m\).
La quantité d’énergie dépensée au total est donnée par la relation :
\(Q = L + S\) \( = 0,58 \times m + \) \(M \times ({T_2} – {T_1})\)
A cette dépense d’énergie, doit s’ajouter le travail mécanique ; d’où le bilan suivant :
Chaleur de combustion des aliments = travail mécanique + chaleur sensible + chaleur latente
b) La calorimétrie indirecte
Elle est basée sur le fait que tout l’oxygène absorbé est utilisée par les cellules pour produire de l’énergie par oxydation des nutriments. On peut à l’aide de divers appareils (spiromètre volumétrique), mesurer le volume de dioxygène absorbé par un animal pendant une période donnée.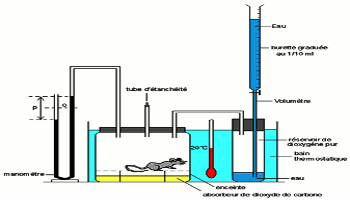
Le coefficient thermique ou énergétique du dioxygène représente la quantité d’énergie libérée par 1L d’oxygène utilisé pour l’oxydation d’un substrat donné.
\({D_E} = {V_{{O_2}}} \times Ct\)
Le coefficient thermique du dioxygène varie en fonction des nutriments consommés : 21Kj pour le glucose, 19 Kj pour les protides et les lipides. Dans une alimentation mixte, le Ct est de 20 Kj
2) Variations des dépenses énergétiques chez les Mammifères
a) Influence des facteurs internes
• L’activité physiologique : le travail digestif, le renouvellement cellulaire et les sécrétions diverses augmentent la consommation de \(O_2\) et donc la dépense d’énergie.
• L’activité physique : la dépense d’énergie chez un organisme homéotherme augmente avec l’activité physique.
• L’âge : La dépense d’énergie est plus élevée chez les jeunes que chez les vieux ou les enfants.
• Le sexe : La dépense énergétique est plus élevée chez le garçon que chez la fille de même masse corporelle.
b) Influence des facteurs externes
La taille de l’animal : plus l’animal est petit, plus sa dépense énergétique est grande : c’est la « loi des tailles ».
La température externe : la dépense énergétique est minimale à la température de neutralité thermique (18-20°C) chez les Mammifères homéothermes. Lorsqu’il fait froid, ces organismes dépensent de l’énergie pour se réchauffer et vice versa
II. Le métabolisme de base
Le métabolisme de base ou métabolisme de repos est la dépense énergétique minimale nécessaire pour assurer le maintien des fonctions vitales.
• Le métabolisme basal se mesure dans des conditions dites« standard ». Ces conditions sont :
• La température de neutralité thermique : c’est la température ambiante pour laquelle la dépense énergétique liée au maintien de la température corporelle est minimale (18°C-20°C). Ceci a pour but d’éliminer la dépense énergétique liée à la régulation de la température interne.
• Le jeun depuis une demi journée : pour éliminer la dépense énergétique liée à la digestion.
• Le repos physique et intellectuel : afin de limiter les activités musculaires et neuronales.
Le métabolisme de base varie avec l’espèce, l’âge, le sexe et l’état physiologique. Il s’exprime en kilojoules par unité de temps (h) et par unité de surface corporelle (\(m^2\)).
\(Mb = \frac{{{V_{{O_2}}} \times Ct}}{{m \times t}}\)
III. L’énergie du fonctionnement cellulaire
1) Nutriments organiques : source d’énergie
Chez les animaux, la digestion aboutit à la simplification moléculaire des aliments en nutriments. Ces nutriments sont utilisés par les cellules pour produire de l’énergie. La mesure de la valeur énergétique des nutriments par combustion dans une bombe calorimétrique montre que :
• 1g de glucide dégage 4 Kcal soit 16,7 Kj
• 1g de protide dégage 4 Kcal soit 16,7 Kj
• 1g de lipide dégage 9 Kcal soit 37,7 Kj
Donc l’énergie qu’utilisent les cellules proviennent des nutriments. Deux phénomènes biologiques sont responsables de la production d’énergie dans les cellules : la respiration et la fermentation, ce sont des réactions de dégradation (réactions cataboliques) qui produisent de l’énergie (réactions exergoniques ou exothermiques).
2) La respiration cellulaire
a) Définition
La respiration est tout processus de dégradation totale des molécules organiques en présence de O2 afin de produire de l’énergie sous forme d’ATP et des déchets exclusivement minéraux (eau et \(CO_2\))
b) Etude quantitative de la respiration
• Le quotient respiratoire (QR)
Le quotient respiratoire est le rapport du volume de \(CO_2\) rejeté au volume de \(O_2\) absorbé; mesuré dans les mêmes conditions de température et de pression.
Le QR varie en fonction du nutriment consommé
\(QR = \frac{{{V_{C{O_2}}}}}{{{V_{{O_2}}}}}\)
• QR = 1 pour les glucides,
• QR = 0,71 pour les lipides,
• QR = 0,83 pour les protides,
• QR = 0,85 pour une alimentation équilibrée (comportant glucides, lipides et protides).
• Glucides (cas du glucose) :
\({C_6}{H_{12}}{O_6} + \) \(6{O_2} \to \) \(6C{O_2} + 6{H_2}O\) \( + 2860Kj\)
– Lipides (cas de la trioléine):
\({C_{57}}{H_{104}}{O_6} + \) \(80{O_2} \to \) \(57C{O_2} + 52{H_2}O\) \( + 35160Kj\)
\(QR = \) \(\frac{{57 \times 22,4}}{{80 \times 22,4}}\) \( = 0,7125\)
– Protides: \(QR\) théorique ≈ 0.85 (valeur obtenue par la méthode de pondération des masses de Berthelot).
NB : Plus le quotient respiratoire est grand, plus la production d’énergie par litre de dioxygène consommée est importante.
On peut donc conclure que le QR renseigne sur la nature des nutriments impliqués dans les phénomènes respiratoires. Dans le cas d’une alimentation équilibrée (comportant glucides, lipides et protides), il est admis conventionnellement que QR = 0.85.
Certains troubles métaboliques comme le diabète peuvent modifier la valeur du QR.
• L’intensité respiratoire (IR)
C’est le volume d’\(O_2\) absorbée ou de \(CO_2\) rejeté par unité de masse de tissu et par unité de temps
\(IR = {V_{{O_2}}}\frac{1}{{t \times m}}\)
\(IR = {V_{C{O_2}}}\frac{1}{{t \times m}}\)
Ou t est donné en heures(h) et m en kilogrammes (kg) et IR en l.h-1.kg-1
N.B : L’IR varie en fonction de l’activité physique, de la température externe chez les endothermes, de l’âge, du sexe et de l’état physiologique. D’une manière générale, l’IR est étroitement liée à la libération d’énergie par l’organisme.
3) Les fermentations
a) Définition
La fermentation est aussi une voie de production d’énergie. Ce sont des oxydations incomplètes de métabolites avec production d’énergie sous forme d’adénosine triphosphate (ATP = Nucléotide constitué de trois molécules d’acide phosphorique). Les déchets produits lors de la fermentation sont totalement ou partiellement organiques.
Certaines fermentations se déroulent en absence d’oxygène (fermentation alcoolique, lactique et butyrique) et d’autres se déroules en présence de dioxygène (fermentation acétique)
b) Les types de fermentation
♦ Fermentation alcoolique ou éthylique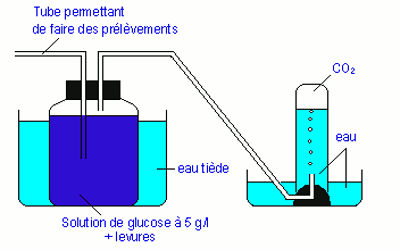
C’est la transformation en milieu anaérobique du glucose en alcool éthylique ou éthanol et en \(CO_2\) cette réaction est catalysée par des enzymes produites par les levures. Ce type de fermentation s’applique dans les brasseries pour la fabrication de la bière ou encore dans les boulangeries pour faire gonfler la pâte.
L’équation de la fermentation alcoolique est :
\({C_6}{H_{12}}{O_6} \to \) \(2C{O_2} + 2{C_2}{H_5}OH\) \( + 138KJ\)
\(\mathop {{C_6}{H_{12}}{O_6}}\limits_{glocuse} \to \) \(2C{O_2} + \) \(2\mathop {{C_2}{H_5}OH}\limits_{éthanol} + \) \(138KJ\)
♦ Fermentation lactique
C’est la transformation anaérobique du lactose en acide lactique. Cette transformation est assurée par les bactéries du genre lactobacille et par les streptocoques.
L’équation de cette transformation est :
\({C_{12}}{H_{22}}{O_{11}} + \) \({H_2}O \to \) \(4C{H_3}CHOH – COOH\) \( + 273KJ\)
Les agents de la fermentation lactique sont des espèces de bactéries lactiques (lactobacilles en bâtonnets et les streptocoques en chaînettes). On a pu démontrer expérimentalement que l’acidification du milieu est fonction du développement des bactéries (graphe)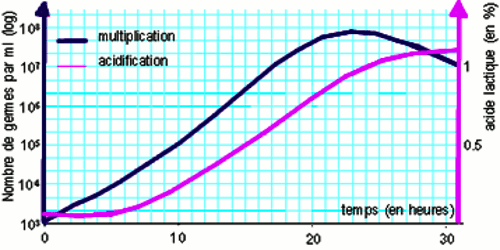
Ce type de fermentation est utilisé pour la fabrication du yaourt.
♦ Fermentation acétique
C’est la transformation de l’éthanol en acide acétique. Cette transformation est assurée par bactéries acétifiantes (Mycodermaaceti) qui utilisent l’énergie produite pour assurer leur métabolisme. L’énergie libérée au cours de ce processus est utilisée par le Mycoderme pour son métabolisme.
L’équation de cette transformation s’écrit :
\({C_2}{H_5}OH + {O_2}\) \( \to C{H_3} – COOH + \) \({H_2}O + E\)
Ce type de fermentation permet la transformation du vin en vinaigre.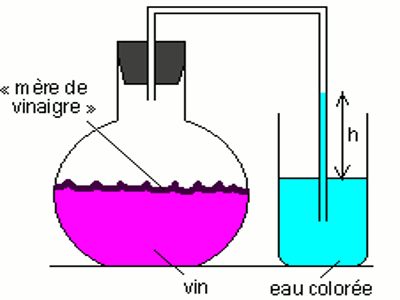
♦ Fermentation butyrique
C’est la transformation anaérobie de l’amidon et de la cellulose en acide butyrique. Cette transformation est catalysée par les enzymes produites par les bactéries du genre Clostridium.
La réaction de cette transformation s’écrit :
\(({C_6}{H_{10}}{O_5})n\) \( + n{H_2}O \to \) \(nC{H_3} – C{H_2} – CH\) \( – COOH + \) \(2nC{O_2} + 2n{H_2}\) \( + E\)
Cette transformation s’applique dans la fabrication du tapioca ou des bâtons de manioc.
IV. Le rendement énergétique
♣ Le rendement énergétique est le rapport de l’énergie mise à la disposition de la cellule sous forme d’ATP par l’énergie potentielle du métabolite.
Sachant que la dégradation complète d’une molécule de glucose (respiration) produit 38 ATP, que la valeur énergétique du glucose est de 2860 Kj et celle de l’ATP est de 31Kj. On peut calculer le rendement énergétique de la respiration (RER).
RER= (38×31/ 2860)x100= 41,18%.
La fermentation par contre est une dégradation incomplète qui produit 2 ATP.
Le rendement énergétique de la fermentation serait donc :
REF=(2×31/ 2860)x100= 2,16%
Le rendement de la respiration est donc largement supérieur à celui de la fermentation, ceci s’explique par le fait que la fermentation est une dégradation incomplète des métabolites.
1) Comparaison de la respiration et de la fermentation
| Points de divergence etre resipi ration et fermen tation | Points communs etre resipira tion et fermen tation | ||
| Elément de compa raison | respiration | fermentation | Dégradent le glucose Passent par la glycolyse Dégagent de la chaleur |
| localisation | Cytosol + mitochondries | cytosol | |
| Oxydation du substrat | complète | Incomplète | |
| milieu | Aérobiose stricte | Anaérobiose | |
| rendement | 38 ATP | 2 ATP |
2) L’ATP : seule source d’énergie directement utilisable par les cellules
L’ATP ou Adénosine Triphosphate est une molécule universellement répandue chez les êtres. Elle renferme dans sa structure un sucre (le ribose), une base azotée (l’adénine) et 3 groupements phosphates formant une courte chaine. En présence de l’ATPase, le phosphate terminal se détache libérant l’ADP (Adénosine Diphosphate), un phosphate inorganique (Pi) et de l’énergie utilisable par les cellules (31Kj).
Dans une cellule, non seulement les réserves d’ATP sont très faibles, mais l’ATP est consommé en permanence. Pour ce fait, les molécules d’ATP sont continuellement synthétisées à partir de l’ADP et du Pi grâce à une enzyme appelée ATPsynthétase : c’est la phosphorylation de l’ATP. Phosphorylation et hydrolyse de l’ATP sont deux réactions couplées
\(ATP + {H_2}O\) \(\rightleftarrows\) \(ADP + Pi\) \( + 31KJ\)
Le blocage de l’hydrolyse de l’ATP par des substances chimiques spécifiques comme le cyanure ou le chloroforme, entraine l’arrêt immédiat de toutes activités cellulaires. L’ATP est donc la seule source d’énergie directement utilisable par les cellules.
V. Les grandes étapes de la synthèse de l’ATP
1) La glycolyse
La glycolyse est la première étape de la respiration et de la fermentation. La glycolyse est un phénomène anaérobique qui se déroule dans le cytoplasme. C’est une suite de 10 réactions qui dégradent une molécule de glucose (C6) en deux molécules d’acides pyruviques ou pyruvates (C3) avec synthèse de deux ATP.
La réaction fondamentale est l’oxydation du glucose en acide pyruvique. Il s’agit d’une déshydrogénation catalysée par une déshydrogénase en présence d’un transporteur d’hydrogène (\(NA{D^ +}\)) qui se trouve alors réduit (\(NADH_2\) ).
L’équation de la glycolyse est :
\({C_6}{H_{12}}{O_6} + \) \(2NA{D^ + } + \) \(2ADP + 2Pi\) \( \to 2C{H_3}COCOOH\) \( + 2NAD{H_2}\) \( + 2ATP\)
2) La respiration : cycle de Krebs et chaîne respiratoire
♣ Décarboxylation oxydative : élimination d’un groupement \(C{O_2}\) d’une molécule sous l’action d’une enzyme, la décarboxylase), qui catalyse l’oxydation de la molécule en présence de coenzyme.
♣ Déshydrogénation : élimination de deux atomes d’hydrogène d’une molécule par une déshydrogénase lors d’une oxydation en l’absence d’O2. Ces atomes sont récupérés par un transporteur oxydé (\({T^ + }\) ou Tox) qui devient réduit \(T{H_2}\) ou Tred), alors que le substrat, réduit devient oxydé.
Coenzyme A : molécule organique dérivant d’une vitamine (groupe B). Le coenzyme A transporte un groupe acétyle \(C{H_3} – CO\) provenant de la dégradation des glucides, lipides et acides aminés dans le hyaloplasme vers la matrice de la mitochondrie où se poursuit la dégradation du radical par le cycle de Krebs.
♣ Acide oxaloacétique \(HOOC – C{H_2}\) \( – CO – COOH\) : acide carboxylique à 4 atomes de carbone qui fixe le groupement acétyle apporté l’acétyl-CoA et donne un acide carboxylique à 6 atomes de carbone, l’acide citrique.
♣ Transporteur d’hydrogène : molécule oxydée de type coenzyme capable de fixer et de transporter deux protons et deux électrons. Le transporteur est alors réduit.
♣ Chaîne respiratoire : chaîne d’oxydoréduction contenue dans la membrane interne de la mitochondrie. Il s’agit d’un ensemble de transporteurs d’hydrogène classés suivant leur potentiel redox croissant. Les électrons parcourant cette chaîne engendrent un champ électrique responsable du pompage, contre un gradient de pH, des protons \({H^ + }\) qui s’accumulent dans l’espace inter membranaire des mitochondries.
Les mitochondries sont les organites cellulaires où se déroulent les réactions proprement dites de la respiration. La matrice mitochondriale riche en enzyme (décarboxylases, déshydrogénases….) est le siège du cycle de Krebs.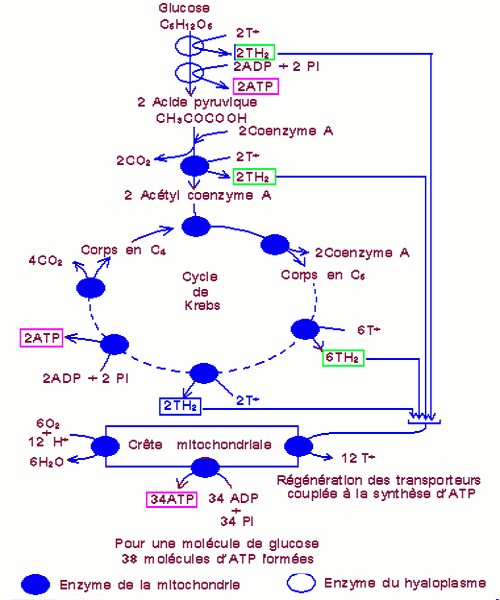
Le cycle de Krebs ou cycle de l’acide citrique est une série de décarboxylations oxydatives, de déshydrogénations, associées à des réductions des transporteurs et à des hydratations.
L’acide pyruvique formé lors de la glycolyse subit une décarboxylation et une déshydrogénation pour donner 2 molécules d’acide acétique (C2). Ces derniers se lient aux coenzymes A pour constituer l’acétylcoenzyme A (acétylCoA). L’acétylCoA s’associe à l’acide oxaloacétique (C4) pour former l’acide citrique (C6). L’acide citrique subit 2 décarboxylations et 4 déshydrogénations pour donner un groupement acétique et l’acide oxaloacétique est régénéré.
Le cycle de Krebs permet donc la libération de dioxyde de carbone, la production d’ATP et la formation des transporteurs réduits \(T{H_2}\) qui sont réoxydés dans la chaîne respiratoire (chaîne d’oxydoréduction) au niveau de la membrane interne de la mitochondrie, ce qui permet ensuite la synthèse d’une grande quantité d’ATP.
Bilan de la respiration
| Différentes étapes | Transporteurs réduits formés | ATP produits |
| glycolyse | 2 NADH2 | 2 ATP |
| Décarboxylation du pyruvate | 2 NADH2 | 0 |
| Cycle de Krebs | 6 NADH2 + 2 FADH2 | 2 ATP |
| Réoxydation au niveau de la chaine respiratoire | 34 ATP |
NB : La réoxydation au niveau de la chaine respiratoire d’une molécule de NADH2 produit 3 molécules d’ATP tandis que celle d’une molécule de FADH2 en produit 2. Au final la dégradation complète d’une molécule de glucose produit 38 molécules d’ATP.
3) Les fermentations
♣ la fermentation lactique
L’acide pyruvique (accepteur final d’hydrogène) issu de la glycolyse est réduit en acide lactique.
Cette réaction est couplée avec la régénération du transporteur d’hydrogène.
Acide pyruvique + \(T{H_2}\) → Acide lactique + \({T^ + }\)
♣ la fermentation alcoolique
L’acide pyruvique est d’abord décarboxylé. Le métabolite qui en résulte (l’acétaldéhyde) est ensuite réduit en éthanol avec régénération du transporteur d’hydrogène.
Acide pyruvique → acétaldéhyde + \(CO_2\)
Acétaldéhyde + \(T{H_2}\) → éthanol + \({T^+}\)
NB : les réactions de fermentations produisent 2 ATP pour une molécule de glucose. Cet ATP n’est produit que lors de la glycolyse.
You may also like

Exercices corrigés Atomistique
Cours d’atomistique Chimie niveau Universitaire [PDF]